In this study an attempt is made to describe ventricular fibrillation with a two-level model. The first, lower level will be discussed in this chapter and in the next as self-sustained activity within a relatively small group of interconnected cells, based upon the results of the single channel analysis of chapter 4. The second, higher level (chapter VIII & chapter IX ) describes the interactions between these groups, based upon the multi channel analysis of chapter 7. The availability of large, digital computers created the possibilty of modelling the electrical activity within the myocardium as the result of interactions between individual cells, instead of approximating this activity by field equations and approximating the solution of these equations by a digital computer. The models intended are thus discrete (contrary to analog) and automatic (contrary to input-output systems), as the myocardium does not react to normal, physiological stimuli during ventricular fibrillation. A "discrete automaton" or simply "automaton" designates a model which has the following features:
A mathematically more rigorous definition of an automaton would be (Barto 1975):
This description leads to the following extension of the definitions given above for the cells:
Implicit in the description above is that x, y, qa and c(i,j) are finite integers. The state qm is real valued, but h(qm,x) will be chosen in such a way, that the number of memorized states per realization is finite. So the polyautomata described in this chapter are finite, uniform, deterministic, Moore-type (discrete time steps with a unit delay between an input and the associated output), autonomous or non-autonomous and static.
Clearly, h(qm,x), C and the begin condition of C remain to be specified for a particular model. In general the evolution of these systems is computationally irreducible to a more simple mathematical formula. Moreover, undecidable problems can arise in the mathematical analysis of such systems (Wolfram 1984). First an attempt has been made to study these models of ventricular fibrillation by lumping all elements together and look for the averaged, overall behaviour. The unsatisfactory results led to mathematical models that allowed for the study of individual elements.
Several of these models have been described.See Bess 1970, Accardi 1972, Anninos 1972 and Burattini 1972. All agree upon the ease with which the networks show reverberations or periodical self-sustained activity.
| variable | value |
|---|---|
| Te | 0 |
| Ta | 1 |
| Tr | 3, 4 or 6 |
| Tm | 65000 |
| h(qm,x) | 1 |
| C | c(i,j) = 0
except for 10 "randomly" chosen j's = i, in such a way that the whole network is connected; |
| M | 1000 |
| begin condition of Q: | begin condition: q(i) = 0,0
except for a number of randomly chosen cells |
In model L1 all cells possess a fixed number of connections with the other cells in the network, all cells have the same probability of being connected with each other and if a cell is connected to an active cell that cell also becomes active. The next step of simulated time the cell is no longer active, but refractory for a fixed number of time steps. Thereafter the cell becomes resting until stimulated. In a sufficiently large network the number of cells that become active may be supposed to be equal to the number of resting cells times the probability that at least one of the connections is active, regarding the total amount of activity at that moment in the network.
The model is started by introducing a certain percentage of activity into the network. A too low or too high percentage of activity does not elicit continuous activity, but the exact borders between persistent activity and transient activity have not been determined. The periodicity is equal to Tr. The reader can verify this easily by changing the parameters of the model LumpedL1. Whether the probability of becoming active is used in a deterministic way to get the actual number of active cells or stochastically by drawing that number from a binomial distribution did not make much difference, so the above mentioned model uses the expectation.
The assumptions in those models are too unrealistic to use these results for anything else than to show that a network of interconnected excitable elements easily exhibits periodical activity, even if the elements themselves do not show periodical activity. For an extensive, mathematical treatment of networks of logical neurones, see Holden 1976.
Based upon the ideas of Waltman a model has been built using a modification of the simulation program Block CSMP of the IBM 1130.
Te 4
Ta 124
F
Tr 148 (K·M + Ta)
Tm 65000
M 1000
h(qm,x) = qm+1
F
d(qm,x) = exp((ln (qm + K·M ) - ln K)/F) with K=90 and F=-0.19
Specially built delay lines were needed for the states: "excitation",
"activity" and "refractoriness". The first three states had fixed delays
of resp. 4, 120 and 24 msec, the last state had a variable delay depending on the overall activity, a maximal refractory period K and a
'influence factor' F (F <= 0). These times and factors were based partly
upon the pictures published by several authors (Sano 1958,
Krinsky 1973, Rushmer 1976, Cranefield 1975) and
partly upon preliminary experiments with the CSMP model to find a stable
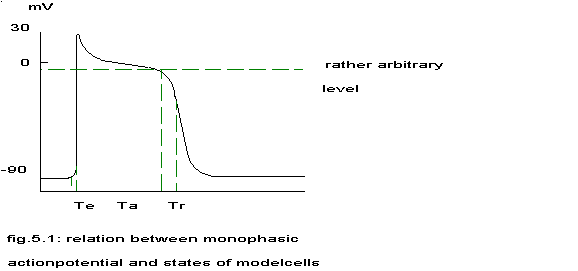
continuous activity pattern. The relationship with the monophasic actionpotential is sketched in the next figure.
|
 |
 |
The known models that describe activity during ventricular fibrillation in terms of coupled elements, do not mean myocardial cells as elements, but - implicitly or explicitly assuming a myocardial syncytium - their elements are needed to discretisize otherwise intractable field equations. See e.g. van Capelle 1980, Foy 1974, Moe 1964, Ritsema van Eck 1972 and Mitchell 1992). At the microscopic level however, the assumption of a syncytium does not work very well, see Spach 1983. In this chapter however the "cells" of the cellular automaton are intended as models of real myocardial cells.
A general theoretical frame to analyze the behaviour of coupled, excitable, non-periodic elements is given by Winfree 1980. Practical considerations like available computer time and memory led to the construction of a model consisting of a three-dimensional array of 1000 cells (10·10·10). The essay of Fozzard 1979 on the conduction of the action potential in the heart establishes the anatomical feature called gap junction or nexus as the correlate for the electrical coupling between cells. The more nexuses, the stronger the coupling, see Plonsey 1974, Kensler 1977, Mann 1977, De Mello 1980, De Mello 1982a and De Mello 1982b. Based upon the anatomical data of Sommer and Johnson (Johnson 1967, Sommer 1979) the number of nexuses between cells was drawn from a binomial distribution, with a probability P and a size of 70. Tissue anisotropy and variation in junctions are sufficient for the necessary spatial variation; the assumption of variation in membrane properties is not necessary (Spach 1983). The cells are considered symmetrical and elongated in the X-direction; they have their nexuses only in the intercalated disks with a strong preference for the X-direction according to the following table:
| p | z-1 | z | z+1 | illustration |
|---|---|---|---|---|
| y-1 | 0.02 | 0.04 | 0.02 |  |
| y | 0.04 | 0.26 | 0.04 | |
| y+1 | 0.02 | 0.04 | 0.02 |
To resolve the
problem of the border (a hypertorus was not considered an appropriate
model) all 488 bordercells were connected to extra cells in a shell
around the threedimensional structure. This shell could either be quiescent or be subjected to different types of activity to simulate the influence of the surrounding tissue on the simulated block. This random
structure of cellconnections was created once and stored in a file for
future parameter studies. The discrete timesteps in the simulations were
supposed to represent 2 ms. In all cases the activity was started by
distributing the cells, as uniformly as possible, random over the total
number of timesteps in the states or phases of the activity cycle. The
type of activity and its stability were investigated for the following
models:
In words: If the number (x) of active nexuses (i.e. the number of connections with neighbour cells in the active state) of a quiescent or relative refractory cell exceeds the threshold (D), this cell enters the
excited state; after 2 steps this cell enters the active state, becomes
refractory 62 steps after excitation and quiescent again after 82 steps.
At threshold values (D) of 1, 5, 10, 15, 20 or 30 all cells were very
regularly active with a repetition period of 164 steps; the total simulated period was 1000 steps. At the thresholds of 15, 20 and 30 a slight
variation in the total number of active cells occurred, but of a random
character. The output of the aggregate as a whole was somewhat noisy, but
no periodicity became apparent. Raising the threshold to 35 extinguished
activity within 328 steps; cells became active at irregular intervals.
The 1000 cells of the model could not be uniformly distributed over
the 82 possible phases at the start of simulation. To check the possibility that the observed variation was caused by the initial irregularity, the lengths of the periods were changed to 3, 76 and 100, so
the proportions did not change too much and the 1000 cells could now be
distributed over 100 phases. At a threshold of 30 this model showed an
exact repetition period of 100 steps in individual cells, but the overall
variation in number of cells that became active was still there. This
variation was too irregular to impose a periodicity on the whole model
and was caused by the fact, that after the initial activation some cells
cannot become excited and/or active as not enough of their neighbours are
active. After a few cycles however all cells in this network ran as fast
as they can, although the threshold of 30 is higher than the average
number of connections between the most strongly connected cells:
0.26·70 = 18.2.
3.1 no input integration, fixed refractory period
f{(Te,qm),x} = Te+1,0
Te 2
Ta 62
Tr 82
Tm 65000
M 1000
h(qm,x) = qm+1
d(qm,x) = D
3.2 input integration, fixed refractory period
The behaviour of real excitable cells, that only become active after
stimulation during a certain time period, was simulated in the following
way: an excited cell will enter the active state only if its number of
active nexuses after Te steps still exceeds the threshold.
f{(Te,qm),x} = 0,0 if x < d(qm,x)
= Te+1,0 if x >= d(qm,x)
The other parameters were not changed and the model behaved at thresholds
of 30 and 31 just as above. At threshold 32 the total activity diminished
slowly during the 1000 simulation steps and the cells fired less regular.
At threshold 33 all activity ceased after 888 steps and at 35 after 425
steps; the firing pattern was totally irregular. Although the aggregate
showed no periodicity in its stable activity at thresholds 30 or 31, the
number of cells that became active at each timestep ranged from 7 to 17
with an average of 12.
3.3 integrated input, relative refractory period
A certain periodicity is measurable outside the model only if the constituent cells should synchronize in such a way, that large groups of
them would have the same phase. The check at the end of the excited state
is apparently not enough for this purpose. At this stage of model building
the quiescent state was replaced by the relative refractory state.
for Tr < qa < Tm: f{(qa,qm),x} = qa+1,h(qm,x) if x < d(qm,x)
= 1,0 if x >= d(qm,x)
h(qm,x) = qm + 1 - F·x/N ( 0 <= F < 1 )
d(qm,x) = exp(-0.22(qm - 21)) + D
N = total number of nexuses of a cell
In words: Cells will enter this state from the absolute refractory state
and leave it for the excited state if their total number of active
nexuses exceeds the threshold. Accomodation (Katz 1966) of the cell
to sub-threshold stimuli is arrived at by subtracting from qm the number
of active nexuses divided by N times a factor (F). The influence of this
factor is sketched in the next table.
| length of states | total # of steps | F | central cell cycle period | total activity of model | |||
|---|---|---|---|---|---|---|---|
| Te | Ta | Tr | mean | SD | |||
| 3 | 76 | 86 | 2000 | 0 | 92 | 0.44 | stable, noisy |
| 3 | 76 | 86 | 2000 | 0.5 | 98 | 2.53 | regular peaks at start, then noisy |
| 3 | 88 | 100 | 2000 | 0.5 | 109 | 0.51 | idem |
| 3 | 88 | 100 | 2000 | 0.75 | 114 | 1.13 | stable, clear double peaks |
| 3 | 88 | 100 | 2000 | 1.0 | 121 | 0.52 | peaks, slow extinction of activity |
| 3 | 88 | 100 | 200 | 1.5 | - | - | irregular low activity and extinction |
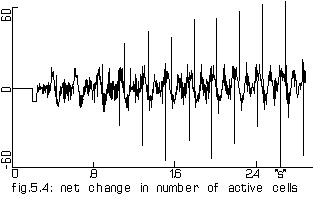 |
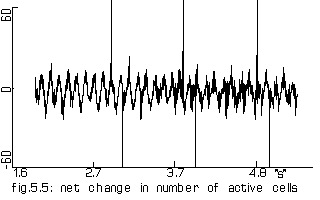
The net change in number of active 'cells' of the model of 1000 cells
when the outer shell was activated every 113 steps (=226 'ms'). This
lasted 85 steps (=170 'ms').
Horizontal axis: simulated time since start of persistent activity Vertical axis: number of cells |
 |
The net change in number of active 'cells' of the model of 1000 cells
when the outer shell was activated with three pulses, each lasting 85
steps (=170 'ms').
Horizontal axis: simulated time since start of persistent activity Vertical axis: number of cells |
This threshold is so low that in the X-direction (the strong coupling) one active cell can activate another, which would result in an excitation front in the X-direction. Analysis of the cells 1,5,5 to 10,5,5 however gave no indication of such a front. The reason for this discrepancy is that approximately a quarter of the cells synchronize in one group and a somewhat larger proportion in another group with a phase shift of half a cycle compared to the first group. The rest of the cells are more or less equally distributed through all phases and their membrane potential variations act as noise to the external electrode. This phenomenon of splitting of the population is described more generally by Winfree 1980 (page 117) for a community of simple, connected clocks, where the aggregate rhythm has two peaks per cycle one-half cycle apart on the average. References to examples and additional analytic models are given by him. A "clock" is defined as a periodical device; its period can be influenced by changing the rate of change of its phase. Winfree also describes in the same book (page 128) how this type of behaviour can also originate in a population of "hourglasses". An "hourglass" being a clock that runs through its cycle only once, this clearly looks like our model of connected cells, as the absence or presence of the pacemaker rhythm of 2000 msec has no influence on the behaviour of the model.
 |
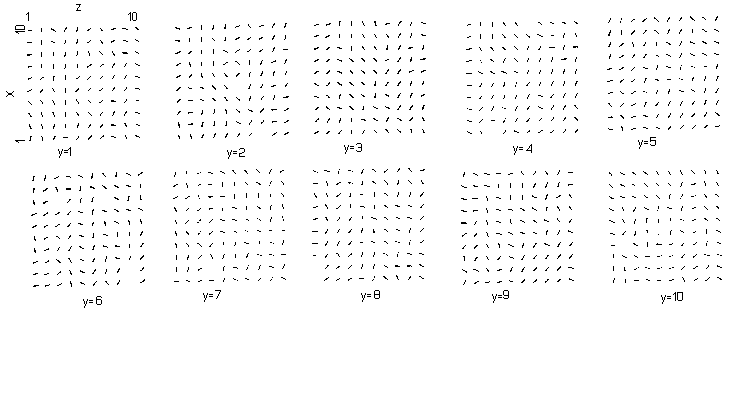 |
The shell of cells covering the model was kept quiescent for the
above mentioned analysis. In order to study the effect of noisy input
from the environment on this small piece of simulated myocard, every time
step this shell could be active with a probability of 0.75. The behaviour
of the central cell did not change, but the total activity did, although
not drastically.
3.6 size considerations
The instabilities of the model are not surprising, if the actual size is
considered. An aggregate of 10 by 10 by 10 cells of the dog myocard will
have dimensions of circa 1.0 by 0.1 by 0.1 mm. Such a small piece of
tissue would stop fibrillation immediately or after several seconds.
(Garrey 1914)
Restrictions on available computing time did not make simulations beyond 12 simulated seconds feasible.
Reduction of the size of the model to 512 cells (8·8·8) led to extinction of the activity within 3000 steps.
| stimulus period | stimulus duration in steps | ||||||
|---|---|---|---|---|---|---|---|
| 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
| 106 | 15 | ||||||
| 105 | 10 | 10 | 15 | 20 | |||
| 104 | 1 | 5,10 | 5,10 | 10 | 15 | 20 | |
| 103 | 1 | 1,5 | 5,10 | 10 | |||
| 102 | 1 | 5 | 5,10 | 10 | |||
| 101 | 1 | ||||||
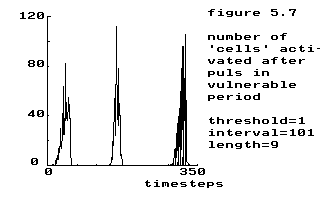 |
threshold = 1
interval = 101 length = 9 |
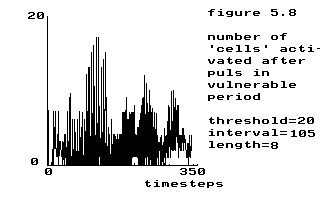 |
threshold = 20
interval = 105 length = 8 |
Foy only states that the wavefront follows a "fairly small, roughly circular path" near the center of his sheet, but Van Capelle and Allessie both indicate a size of approximately 5 mm for the diameter of this circle, around which one or more wavefronts spiral. Winfree (1980) page 310 states in his treatise of an involute spiral wavefront: "the thinness required to justify a two-dimensional description is about the diameter of the troublesome central disk of the involute approximation". The ventricle is much thicker than this 'central disk', so very complicated patterns might develop. Whether the complex periodical pattern of fig. 5.6 can be described as one of Winfree's three-dimensional rotors has not been investigated.(Winfree 1980)
The splitting of the population of cells with regard to their phases can be seen in the light of the theory of Winfree on networks of simple clocks as mentioned in paragraph 4.5. See Winfree 1980.
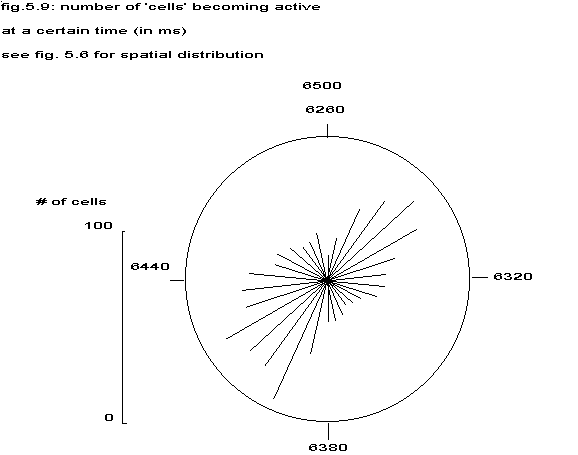 |
see fig. 5.6 for spatial distribution |
The fact that the model exhibits persistent, periodical activity as an oscillator, although the constituent elements are not periodically active (at least not in the reported order of magnitude) is by no means a novel finding. The topological model is written in the language of cellular automata and these can behave as an oscillator (Merzenich 1974). More appropriate to the next level of modeling is that stimulations of the model made clear that the model as a whole can be described as a relaxation oscillator, although no attempt has been made to find the set of differential equations that give the same output as the model. However, in chapter VIII formula's will be given for an oscillator which shows the same overall behaviour as the topological model.
The reader can verify most of the above mentioned results with the program Topolt.